

Introduction
Rice is one of the world’s most widely eaten and most productive cereals. Over the past decade archaeobotanical research based at the UCL Institute of Archaeology and involving a wide range of collaborators in east, south, and southeast Asia has been seeking new insights into the processes by which rice evolved and rice cultivation systems evolved, and how these relate to cultural change and long-distance cultural relationships across Monsoon Asia. While this research began with critical reflections and debate surrounding the initial cultivation of rice in China (Fuller, Harvey and Qin 2007) and India (Fuller 2006), it has expanded into better documenting the evidence for morphological change during domestication (Fuller et al. 2009), the characterisation of variable wet and dry ecologies of early rice cultivation (Fuller and Qin 2009; Fuller and Weisskopf 2010; Weisskopf et al. 2014), and to tracking the spread of rice through the compilation and analysis of a large database of archaeobotanical evidence (Silva et al. 2015). We have completed more than six years of research supported by major NERC (Natural Environment Research Council) grants (Grant NE/G005540/1 “The Identification of Arable Rice Systems in Prehistory” (2009–2012); Grant NE/K003402/1 “The impact of evolving rice systems from China to Southeast Asia” (2013–2016)), developing and deploying new methods for establishing the past ecology of rice cultivation and its potential impact not just on social change, but also on the global environment through methane emissions produced by methanogenic bacteria that thrive in the warm stagnant water of paddyfields (Fuller et al. 2011). The time is right for assessing what we now know and which hypotheses require further archaeobotanical testing. In the present contribution we reflect on some of the key questions about the diversification of rice as it emerged and spread as a major crop around Monsoon Asia.
Rice has long been important for large areas of Asia, but not all of this region has been equally subjected to archaeobotanical sampling. Based on a database of archaeological evidence for rice from across 400 sites (Silva et al. 2015), we can visualize coverage of rice archaeobotany on a heat map (Fig. 1). This shows in hotter (redder) shades areas with a large amount of archaeobotanical evidence for rice, and in lighter shades areas that lack data. This highlights how limited the data are from southern China, northern and Island Southeast Asia, Myanmar and southeast India. Studies carried out by the authors and collaborators in recent years have included archaeobotanical assemblages from northwest Pakistan, the Middle Ganges valley, central coastal Odisha, interior Tamil Nadu and northern Sri Lanka, Thailand, south Vietnam and the Lower Yangtze region and central China (mainly Henan province). In the next few years of the Early Rice Project we aim to tackle some of the key gap zones, indicated in Figure 1 by pink circles. While some conclusions must be tempered by acknowledging gaps in the evidence, especially between northeastern India, Myanmar and southern China, there are many reliable conclusions that we can make from the available archaeobotanical evidence, and from a critical reading of recent advances in rice genetics.
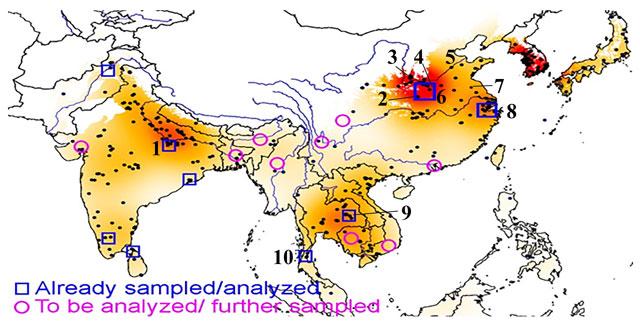
Heat map showing regions with more archaeobotanical evidence for rice, for all periods (the more data, the darker the red). Selected sites/site groups numbered: 1. Mahagara and Koldihwa, 2. Baligang, 3. Sushui valley, 4. Chengyao, 5. Houli culture area, 6. Jiahu. 7. Shushanji, 8. Lower Yangtze region (Kuahuqiao, Tianluoshan, Majiabang, Caoxieshan, Xiaodouli, Maoshan, Liangzhu), 9. Ban Non Wat and Non Ban Jak, 10. Khao Sam Kaeo (Heat map prepared by F. Silva).
When and how was rice domesticated?
In order to understand the origins of rice, it is necessary to recognize that there is more than one kind of rice. Study of genetics has made it clear that the two traditional subspecies of japonica and indica are quite distant, and there is a third lineage of rice – the aus rices – that today are at their most diverse in northeastern India and Bangladesh (Schatz et al. 2014; Travis et al. 2015; Civáň et al. 2015). Modern genetics also indicates that despite the deep divergence between indica and japonica these subspecies share several mutations that have been selected for during domestication, which in turn implies early hybridization between these two cultivated lines; it is generally inferred that many domestication traits evolved first in japonica, then were transferred to a proto-indica through hybridization under cultivation (Fuller and Qin 2009; Gross and Zhao 2014). The aus rices, which until 2005 were simply assumed to be part of the indica subspecies, could represent a truly independent origin, but more work is needed, especially archaeobotanical research in the major gap in current evidence (Fig. 1).
In recent years an increase in systematic archaeobotanical sampling has begun to make possible the documentation of rice domestication in terms of the evolution of morphological domestication traits, the development of arable habitats, and the shift in reliance from wild gathered foods to cultivated rice. Domestication of rice, as with other cereals, involved evolution of non-shattering panicles, i.e. loss of wild type seed dispersal: this has now been increasingly documented since 2006 by the recovery and study of rice spikelet bases (the base of the rice ‘flower’) (Fig. 2). This has been an important methodological development, with some 27 spikelet bases identified archaeologically prior to 2006, and now 10,000s have been documented in China, Southeast Asia and India (Fig. 3). In the Lower Yangtze region these indicate a chronologically protracted evolutionary process over 3000–4000 years (Fuller et al. 2009; Fuller et al. 2014), reaching the domesticated non-shattering state by ca. 3800 BC. In the Middle Yangtze fewer such data are available, but evidence from the Han River (the Baligang site which is located on the Bai River, a small tributary of the Han River), a northerly tributary of the Middle Yangtze, indicates that largely non-shattering (>80%) rice was already established by 6300 BC (Deng et al. 2015). This indicates that selection for this domestication trait started earlier in this region than in the Lower Yangtze, but it also suggests a separate process from the Lower Yangtze. Logically, if there had been a dispersal of rice cultivars from the Middle Yangtze between 6000 and 4000 BC, it would have brought many more domesticated forms than are actually found at sites like Kuahuqiao or Tianluoshan. Instead the slow change documented in the Lower Yangtze indicates a local process of rice evolution. Recent genetic studies have revealed that there are multiple genetic mutations that contribute to non-shattering in modern domesticated rice (Htun et al. 2014), and it is conceivable that initially these different genes evolved in different parts of the Yangtze basin as part of separate domestication processes. Other regions of eastern China that have produced early evidence for rice lack evidence for whether or not such rice was evolving as non-shattering, which can only take place under cultivation, or might have been collected wild. Further archaeobotanical evidence is therefore needed. Such regional cultures with rice consumption in the 7th M. BC, include Jiahu in the Upper Huai river (Zhao 2010), Shushanji in the Lower Huai River basin (Nanjing Museum 2016), and the Houli culture of Shandong Province (Jin et al. 2014).
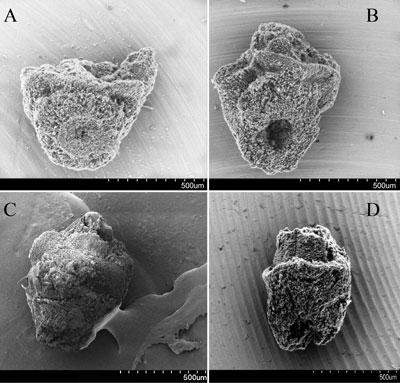
Examples of archaeological spikelet bases of rice, including morphologically wild types (A), domesticated types (B, D) a green-harvested, immature type (C), which still has attached apex of rachilla (the axis of a spikelet). A, B, are Longshan period Chengyao, Henan, China; (C) is from Tianluoshan, Zhejiang China, and (D) from Late Neolithic Mahagara, India. Scale bar 500 microns in each case. (SEM images by DQFuller).
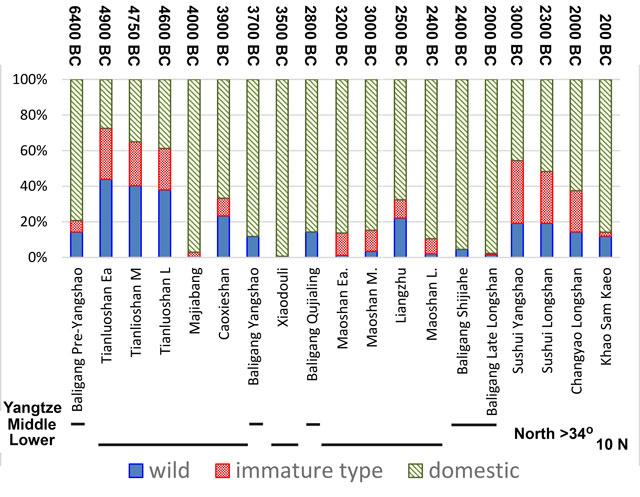
A comparison of the proportions of rice types for various sites and phases in China and Iron Age Thailand, as classified by spikelet bases, separating shattering (wild or weedy), non-shattering (domesticated) and green-harvested (immature). This chart summarizes data on 35,309 identified archaeological spikelet bases studied since 2006 (Chart prepared by DQF).
Spikelet bases have begun to contribute to documenting the evolution of rice in India as well. The Neolithic site of Mahagara, where macro-remains and phytoliths document mixed agriculture based on rice, other cereals and pulses since the early 2nd M. BC (Harvey and Fuller 2005), has also produced, as a result of re-examination, a dozen spikelet bases that are predominantly domesticated (Fig. 2D). This indicates domesticated rice, presumably of the indica subspecies, was established in the Middle Ganges valley by 1600 BC at the latest. Prior to this period there is little firm evidence for the extent to which rice was domesticated or wild in South Asia, due to a lack of systematic recovery of spikelet bases. Nevertheless, these limited data are congruent with the idea that indica rice arose through hybridization between local unimproved cultivars that were still morphologically wild (“proto-indica”) and introduced, fully domesticated subspecies japonica.
Another aspect of evolution during domestication was an increase in grain size, but in particular grain girth (breadth and thickness) not length. Change in this trait is also increasingly evident from regional sequences of archaeobotanical data and indicates some variation in separate regional trajectories. The data from Baligang, despite being predominantly non-shattering, consists of thinner grains more like wild rices and not comparable with the fatter rices in the Lower Yangtze that accompanied the rise of non-shattering (Deng et al. 2015). This indicates that grain size evolved more slowly in the Middle Yangtze, where non-shattering evolved more quickly. This also indicates that the trajectories of domestication in the Middle and Lower Yangtze regions were separate and different, with earlier evolution of non-shattering in one region and grain size in the other. In northern India grain size increase took place over the second and first millennia BC and indicates that selection for bigger grains occurred only after a domesticated indica had evolved through local hybridization with introduced japonica (Fuller et al. 2014). The evolution of larger-grained, and higher yielding indica accompanied the growth of population, indicated by more permanent habitation sites and increasing size of larger sites over the course of the Chalcolithic and Iron Age periods throughout the Ganges plains between 1500 and 500 BC, with clear urbanism by the end of this sequence (Conningham 2005). This was also the period when rice became more widespread throughout the Indian subcontinent (Fuller and Qin 2009; Silva et al. 2015).
When did temperate-adapted rices evolve?
Within the evolution of the japonica subspecies, there was another early subdivision between a temperate and a tropical branch, which evolved into different regional adaptations after domestication (Civáň et al. 2015). Temperate japonica rice was particularly important in taking rice to high latitudes, where it became associated historically with important rice production in central and northeast China, Korea and Japan. One of the key ways in which this was possible was by evolution of strong linkage between rice and the summer season, when it is warm enough for rice to grow effectively. This occurred through a genetic change which made temperate japonica highly sensitive to day length, with flowering and grain production brought on by the shortening day lengths as summer progresses. This facilitated rices being planted in the spring, prior to the start of summer, when rain and temperatures are better for rice growth, while forcing flowers and grain production before the colder weather of autumn. While wild rices, and tropical japonica, have variable responses to day length, and often long growing seasons (of around six months) temperate japonica has a shorter growing season and consistently timed flowering towards the end of summer. This is controlled by a distinct mutation at the gene locus known as DTH-A4, which must have evolved after domestication as it is absent from wild rice populations (Wu et al. 2013). This mutation leads to higher levels of grain filling under northerly temperate conditions. Temperate japonica rice also tends to be characterized by shorter, but plump grains, which have consistently lower Length to Width ratio (Kitano et al. 1993). While lower Length to Width ratios characterize most varieties of the subspecies japonica, compared to higher ratios and variability in subspecies indica (Castillo et al. 2016), those to the north tend towards the lower end of japonica ratios.
Archaeobotanically we get two lines of evidence which hint at the evolutionary origins of temperate japonica. First, early assemblages of rice from northern China (in or near the Yellow River basin), when spikelet bases have been recorded, have very high proportions of immature spikelet bases (see Figure 3: Sushui Yangshao and Longshan, Chengyao). This suggests that these early northern rices had inadequate maturation time to fill most of their grains, with higher immaturity levels (>20%) than even semi-wild populations undergoing initial domestication. This is to be expected in rice that has one of the wild type gene variants (allele) (DTH-A2), which is also common in the domesticated rices of southern China; they only fill about 60% of spikelets when grown experimentally under northern conditions (Wu et al. 2013). By contrast, the mutant DTH-A4 fills over 80% of its grains under the same conditions. This means that once DTH-A4 mutants occurred they have around 25% more offspring per plant. This on its own could lead to rapid natural selection for this genotype. Of course, if early farmers noticed some plants were productive, and preferentially planted the seeds of those, then the selection process for DTH-A4 would be even more rapid. Indeed we might expect this gene to become fixed in northern rice populations in just a few human generations, even less than a century—this rapid fixation of post-domestication improvements in crops is expected to be very rapid in contrast to much more protracted evolution of initial domestication traits (Purugganan and Fuller 2009). The archaeobotanical evidence so far suggests that DTH-A4 was selected for only after the Longshan period, i.e. more recently than 4000 years ago. Another gene that contributed to this northern adaptation is Ghd-7-2 which promotes increased branching of stems and panicles under the longer days of summer (Xue et al. 2008). The evolution of genes connected to productivity under temperate summer conditions may well be related to the subsequent dispersal of rice to other northern regions such as the Korean Peninsula (ca. 1500 BC) and Japan (after 1000 BC) (Silva et al. 2015; Fuller et al. 2011).
Another characteristic of temperate japonica, short grains, may have evolved earlier. Among the earliest assemblages of rice in the Yellow River basin are those from Nanjiaokou, which are directly dated to just after 4000 BC. These grains are wide, keeping with a domesticated form, but very short, and indeed many of the reported metrics on Yangshao and Longshan rices suggests that they are on average short (often close to 4mm long, when found charred) (Qin and Fuller 2009; Liu et al. 2007). Thus we can conclude that the evolution of temperate japonica began as soon as it moved north, but intensified after 4000 years ago, around the start of the Bronze Age, when new mutations made northern rices more productive. It was only after this time that rice became a really important part of subsistence economies in north eastern China, Korea and Japan as it has been historically.
What was the ecology of early rice cultivation in different regions?
One of the key axes of variation in rice is the ecology under which it is cultivated. This ranges from deep water, permanently wet systems, to irrigated systems that are flooded through most of the life cycle, to systems that are upland and dependent only on rainfall (Fig. 4). In others words there is a spectrum from wet to dry (Weisskopf et al. 2014; Fuller et al. 2011). In some cases upland rice may be grown in slash-and-burn systems. One of the aims of our research has been to establish indices in the archaeobotanical data to separate drier from wetter cultivation ecologies. This is important in two ways. First, in socioeconomic terms dry rice tends to require less labour input and to be less productive. Rainfed rice may yield from half a metric ton to 1.3 tons per hectare under traditional cultivation, while wet rice systems traditionally yield 2–2.5 tons per hectare. This has obvious implications both for the support of growing population densities in the past, but also for the labour demands of agriculture. The historical record of Southeast Asia indicates that the political dynamics of the past millennium have involved interaction between lowland states, supported by large populations of productive wet rice agriculture, and peripheral hill cultivators that were mostly outside of state control and who practiced lower intensity rainfed farming of rice and other hill crops (Scott 2009). The less work and fewer tax responsibilities of upland rice cultivation may often be more attractive to farmers than the hard work of wet rice. Thus we can ask the questions: when and under what circumstances did wet rice agriculture emerge in Southeast Asia. A second important aspect of wet rice is that these ecosystems produce methane, a significant greenhouse gas that contributes to global warming. Many palaeoclimatic records suggest that methane levels have diverged from the expected natural interglacial levels and increased over the past 5000 years, and that one of the contributing factors is likely to have been the expansion of wet rice farming in Asia (Fuller et al. 2011; Ruddiman et al. 2016). Thus one of the key aims of our Early Rice Project is also to use our growing archaeobotanical database to better infer the extent of the wet rice area in later prehistory and how this contributed to global climate change.

Examples of differing rice cultivation ecologies, clockwise from top left: deep water rice (Odisha, India); upland slash-and-burn rainfed (dry) rice (Laos); dry rainfed rice (Maharashtra, India); lowland irrigated/flooded wet rice (Odisha, India) (Photos DQF/ARW/DQF/DQF).
One of the methods we have developed for inferring whether past rice was wet or dry is to look at the statistical composition of phytolith assemblages of associated grasses on sites with rice cultivation. Based on the principle that most of the grass phytoliths in archaeological middens come from the waste of crop processing and therefore reflect the ecologies of rice fields, we can compare the proportion of phytoliths from different parts of the grass plants (Weisskopf et al. 2015a). While it may not be possible to identify most grass phytoliths to the genus or species from which they come, it is possible to separate those from leaf surfaces, versus pores, leaf interiors or husks. Some of these, such as the typical short cells of the rice leaf surface, are what we call “fixed” types in that they normally always silicify and produce phytoliths. Other cells that perform specific functions, like opening/closing pores, stop functioning when they silicify, and thus they only silicify if the plant has an excess of silica to deposit, and usually later in the life of the plant. The latter we term “sensitive” as they can be expected to form phytoliths under situations of excessive evapotranspiration. Given that all rice is grown in hot climates, the main variable in how much evapotranspiration there is should be water availability, and thus more sensitive forms turn into phytoliths in wetter fields. We have tested this method on phytoliths collected from the soils of modern rice stands, and have also shown logical changes from wetter, to somewhat dry, to wet again in the archaeological sequence of the Lower Yangtze (Weisskopf et al. 2015a).
We have now applied this method across a large number of sites in China, India and in Southeast Asia (Fig. 5). What these data indicate is that early rice cultivation in both the Middle Yangtze region (the site of Baligang) and the Lower Yangtze region (sites of Tianluoshan, Maoshan) was very wet; indeed ratios of >2 (twice as many water sensitive phytoliths compared to fixed silica forming types) are what we find in the wild ancestors of rice that are native to marshes. Interestingly, however, there had been a transition towards somewhat drier cultivation systems, seen in the Lower Yangtze at Caoxieshan (ca. 3900 BC). We have attributed this shift to cultivation in small fields with tight control of flooding and then draining that helped to increase grain production in early rice, which still had ancestral proclivity to perennial growth and lower grain production (Weisskopf et al. 2015a). Through the sequence at Baligang we also see a period when cultivation systems became drier although it is less clear why that was (Weisskopf et al. 2015b). In contrast to the generally wet ecology of early rice in China, Neolithic sites from northern India in the Ganges Basin (Mahagara and Koldihwa) indicate very dry conditions, consistent with cultivation on the plains raised well above the river flood plain, supplied only by monsoon rainfall and without methods of water control. The site of Non Ban Jak, a later Iron Age site from northeast Thailand, represents wet irrigated rice, and is associated with a period in which canals and moats were dug to help manage water, and a period in which ploughing with water buffalo seems to have been established (Higham et al. 2014).
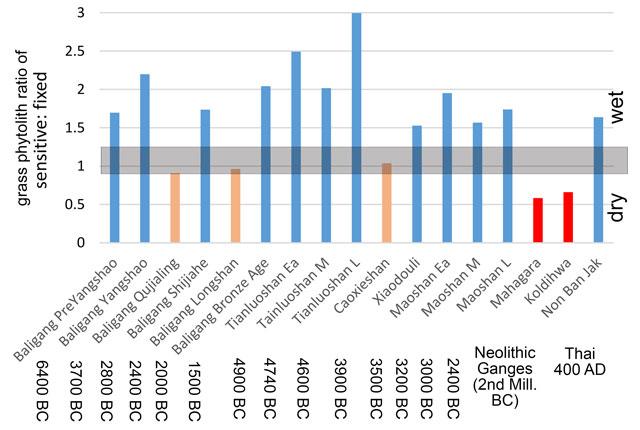
The sensitive: fixed grass phytolith ratio for various sites and phases (averaged across multiple samples), indicating in blue those that are inferred to represent wet field ecology, in red those that are dry field ecology and those in orange that are somewhat dry or intermediate. Phytolith analyses by ARW. Based on 138 samples and at least 300 single cells counted per sample (Chart prepared by DQF).
Much of the rice cultivation in the plains of Southeast Asia today can be regarded as irrigated and wet, in as much as water is retained in the fields by bunding, and water levels can be topped up from various tanks, canals or rivers. But this appears not to have been true at the beginnings of agriculture in the region. We have seen that the phytolith index from Non Ban Jak from the 3rd or 4th century AD was wet. At earlier sites, however, extending back through the 1st and 2nd Millennia BC, we have indications that rice cultivation was in rainfed, dry systems (Castillo 2011; Castillo et al. 2016).
Another approach to reconstructing how rice was cultivated is from the seeds of weeds that were harvested alongside rice. Evidence of early weeds in India indicates a predominance of dry tolerant rather than wet-associated species (Fuller and Qin 2009). In a sequence of archaeobotanical assemblages from the Phimai region in northeast Thailand, the seed assemblages include a large number and range of weed taxa. When these taxa are divided into wet versus dry ecologies (and some that are indeterminate, as they are reported from both), based on modern agricultural studies, it can be seen that earlier phases, represented by Ban Non Wat are dominated by the seeds from dry species, and the diversity of dry species also tends to be greater than those of indeterminate or wet taxa. After the 5th century BC, Ban Non Wat assemblages appear wetter, with more wet taxa, roughly equal number of seeds of wet versus dry indicators and a very large presence of indeterminates. This suggests that the fields, or at least some of them, were becoming wetter. Later, at the nearby site of Non Ban Jak the predominance of a wet field ecology is clear (Higham et al. 2014). Thus we conclude that in the final centuries BC to the early centuries AD rice cultivation was intensified through irrigation and labour investment in water control.
One likely reason why cultivation of dry rice was earlier in Southeast Asia is that it requires much less work. It needs no embanked fields or canals, and less field maintenance. Cultivation is little different from any other seed crop, such as foxtail millet (Setaria italica), which was also cultivated in Thailand and Vietnam from the beginnings of grain-based agriculture around 2400 BC. One of the curious aspects of the early history of rice is how long it took to appear south of the Yangtze basin: with the earliest rice in the Pearl River of southern China dating from ca. 2500 BC, and similar ages for the establishment of rice in the Upper Yangtze region in Yunnan or on the coastal plains of the island of Taiwan (Silva et al. 2015). It can be suggested that the delay was partly about waiting until less intensive dry rice systems, and upland rice varieties, had evolved from the original wet rices of the Yangtze. The societies of southern China were perhaps not ready or willing to expend the high levels of labour needed for wet rice, but once less intensive dry cropping became possible rice agriculture readily dispersed (Fuller and Qin 2009). Dry rice was also suited to a wider range of landscapes, whereas wet rice would have been more restricted to lowlands and floodplains. This has the further implication that the spread of rice farming was not merely the result of rising population densities in the core areas of early cultivation, but was also conditioned by the ease with which low intensity farming could be taken to a wider range of frontier conditions.
The cultural and environmental legacies of rice
Rice has the potential, when grown in wet fields, to be highly productive—two or three times as productive, or even more, than most other cereals. As such, it is not surprising that many states in Asia, from Japan to Southeast Asia to Sri Lanka, have relied to a large degree on the cultivation of wet rice to support their urbanism, trade, armies, etc. Nevertheless, it is also clear that the mere presence of rice did not inevitably lead to higher populations and formation of states, for in many cases rice was much less productive, for example when it was dry cropped only, or before it had evolved adaptations to more northerly environments. In this regard certain contrasts are instructive: early state formation in the Yellow River basin in China, taking place from the end of the Longshan period (ca. 2000 BC) was based predominantly on foxtail millet, with rice playing only a minor subsistence role. As we have argued above this can be explained in part by the delay between the introduction of rice to northern areas and its evolving adaptations to higher productivity (grain filling) under northern conditions. In the context of Southeast Asia, early rice cultivation was based on lower productivity, and lower labour investment rainfed production, with intensification into wet rice systems taking place around 2000 years ago, the same era which sees clear emergence of larger habitation sites, and beginnings of what were to develop into cities and states in northern Thailand. Once established wet rice did indeed support dense populations and growing hierarchical systems.
The wet rice systems that supported examples of state formation also contributed to increasing transformation of the environment. Locally this required control of water—canals, embankments, levelling of fields—meanwhile, globally, the expansion of wet rice contributed to rising trends in global methane that have contributed to global climate warming probably for several millennia (Ruddiman et al. 2016). Other human activities, like deforestation and the spread of livestock must be taken into account in global methane emissions prior to 2000–3000 years ago, but rice was likely to have been important. Our recent efforts to improve the archaeobotany of rice, not merely by collecting rice from more periods and places, but also by employing new methodologies for inferring how rice was cultivated and when it was truly domesticated, have contributed to understanding rice as a key component in social and ecological systems as they evolved and diversified in tropical Asia over the course of the Holocene. Over the next few years we hope to further refine our methodologies in analysing rice and weed seed assemblages, and phytolith assemblages, and extend this work to more regions in Asia and thus help fill in the geographical and chronological gaps in the archaeology of rice.